|
NOTE
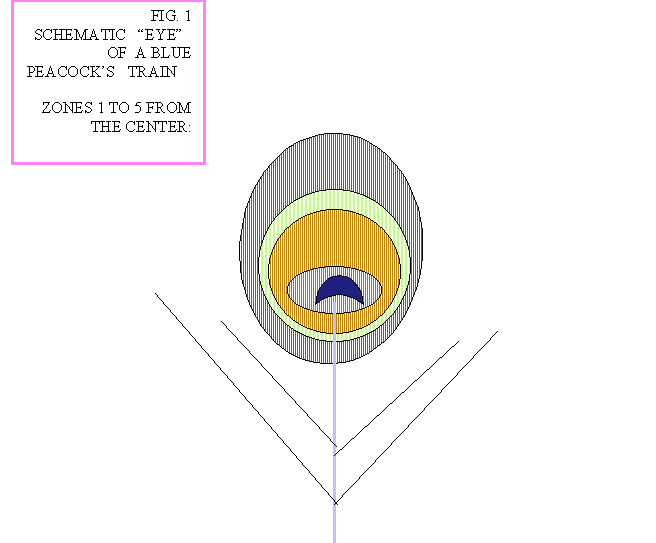
In the following you may find the Article about the physical characteristics of the "eye" of
the Peacock train, as was last revised in March 2000. At the bottom of this article you may find Fig.1 that gives
a schematic picture of the "eye" with the 5 zones, numbered from the center, that are mentioned along the
article. The article itself may requier additional additing. But even in its status now, one may learn the main
intresting ideas of the article. Improvements may be done in the near future. In the next version we
may take in account the notes of the readers, including your notes, if you may kindly use the "Contuct Us" button
to send them to us. Thank you and enjoy the ideas of the Article.
______________________________________________________________________
23 March, 2000
Israel Fried
Physical explanations to the "eye" in the train of the blue peacock (Pavo cristatus L)
Abstract
I suggest physical explanation to the "eye" in the train of the blue peacock, i.e. the male of the
Indian Peafowl (Pavo cristatus L). Generally, a bird's tail is used for stabilizing weight balance and aerodynamic forces
when flying and when perching on a branch. The genus of the birds Pavo pheasants are those with the "eye" like pattern on
the edge of their long tail (train). The source of those pheasants is in the jungles of SouthEast Asia where there are strong
winds due to the monsoons all over the year. The blue peacock was used to roost and foraging for food at the top of the taller
trees, probably in order to bring high quality food to the females and young waiting on the ground or on the low branches.
The long train belongs to the males only.
I show that the "eye" might function as an automatic controller against strong winds. I suggest that
the control mechanism be by enabling a separation of the barbules in those "eyes" by the right amount under all kinds of winds.
This include turbulent winds coming from many directions in a variety of strengths, while the winds may be changed abruptly
locally on each separate feather, as might be the case at the top of the tall trees in the monsoons jungles. I recall works
of others, which show that the colors in the "eye" are due to interference from melanin lattices with specific spacing between
the layers. I suggest a connection between the quantity and density of the melanin layers and the mechanical properties of
the barbules, which allow the opening of the barbules in each colored zone under specific levels of wind intensity.
1. Introduction
There are a many works in biomechanics regarding the mechanical functions of animals and plants.
See e.g. recent reviews about the silent flight of the owl [Sefile 1999], about physical dimensionless numbers in organisms
[Vogel 1998] and an old but very interesting book regarding structure form and movement in animals [Hertel, 1966]. In spite
of all those works, the meanings of colors in animals and plants are regarded yet as due to social communicative reasons.
See e.g. recent review about density dependent warning coloration in Acridadae [Sword 1999]. According to my new theory "ZooPhysics"
one may suggest primary physical explanations to the shapes, colors and patterns in animals. I also develop another new theory
"PlantoPhysics" for explaining the same in plants. In ZooPhysics we ask how the colors and shapes of each part of the animal
helps it to reach its special food while it was in the specific habitat conditions in the wild source homeland. I regard those
functions of the shapes and colors of animals, as the primary functions. The other functions, such as protecting against enemies,
communicating with the other sex, camouflage, etc., are regarded in ZooPhysics as secondary functions.
The wonderful "eye" pattern (ocellus) in the feather's train of the "blue peacock", i.e. the male
of the Indian Peafowl (Pavo cristatus Linnaeus), is a very good example of a biological subject which may be treated from
physical aspects. However, it is regarded mainly as an instrument to impress the peahen [Petrie 1994], as well as other communicating
functions of the peacock with its surroundings. Therefore, I choose this subject as the first article in ZooPhysics.
The structure and form of the "eye" of a blue peacock's feather and other birds were investigated
intensively [Durrer, 1965, Lucas, 1972; Welty, 1982]. Pictures and special behaviors of blue peacocks and peahens are described
in many bird's guides, e.g. [Whistler, 1941; Gilliard, 1967; Henry, 1971; Johnsgard 1986]. Nowadays, one may find nice pictures
of blue peacock's train in many INTERNET sits using search engines with the key words e.g. "Pavo cristatus", "blue peacock",
etc. One may see also the movements of the blue peacock in videocassettes and TV programs on animals, etc.
Here I emphasize the differences between the foraging behavior of the peacock and the peahen in their
source homeland wild habitat. Then I describe the physical functions of the long train of the male and the suggested physical
function of the "eye. Thereafter I talk about the specific structures of the barbules in each color zone of the "eye". I show
how those structures may help the male to stand against strong winds, while foraging special food at the top of the tallest
trees in the monsoon jungles of India and Seri-Lanka (Ceylon). But before all that, I wish to start with description of very
simple experiments, which one may try in it's home, just to indicate the extraordinary physical functions of the marvelous
feather of a blue peacock's train.
2. Simple Experiments
Let us discuss several simple experiments revealing some of the mechanical characteristics of a typical
long feather from the train of the blue peacock. (One may note that there are like feathers of the other species of the Pavo
genus, the Green Peafowl, but their colors are somewhat different). Holding vertically such a feather, e.g. of length 80 cm,
reveals it's extraordinary lightness in regarding to it's length. About the lightness of birds feathers, see e.g. pp.51-55
of [Hertel 1966]. When I hold the feather in its base and move it horizontally from side to side, I feel that the aerodynamic
resistance on the ventral of the feather is slightly stronger relative to that on the dorsal side, i.e. where the colors of
the eye" are seen. Holding the base of the feather by two fingers" while keeping it horizontally, and move it up and down
I see the slow falling of the feather's edge due to aerodynamic resistance. All these experiments show the marvelous aerodynamic
features of that feather. Such features may explain why the peacock moves its train so easily although there might be 150
or more feathers (at various lengths) in a train. Now let us consider the "eye" itself at the palm-like in the edge of the
long feather of a blue peacock's train. There are several color zones in that "eye". As noted in many references, these are
physical colors, which depend on the angle of sight and the spectrum of light reaching the "eye". While facing such a typical
"eye" in a clear daylight one may recognize several zones of colors. Here we shall talk about the following main zones, from
the center of the "eye" outwards (see Fig 1):
Zone 1: DarkBlue, heart shaped at the center.
Zone 2: Turquoise, pressed circle shaped.
Zone 3: RedBrown egg shaped.
Zone 4: GoldenYellow, ring shaped.
Zone 5: Green-Red, wide area up to the edge of the "eye.
The colors in these zones may be changed due to viewing the "eye from different angels.
The first hint I got to a physical function of a peacock's "eye was while I was trying to separate
gently, by two fingers, two barbs in the "eye". I recognized interesting behaviors of the colored zones. The separation along
the RedBrown zone 3 goes relatively slowly. The separation along the turquoise zone 2 goes slightly faster. But when I reached
the beginning of the DarkBlue zone 1, the two barbs were separated spontaneously all along the way. These simple experiments
may suggest that there is a specific mechanical function to each colored zone in the "eye.
Since the train may be regarded as an aerodynamic stabilizer, it was important to search the "eye"
under different wind conditions. The simplest qualitative experiment is by blowing of wind from one's mouth toward the "eye".
One may realize that different forces of airflow are required in order to separate the barbs of different colors. The closer
the shade to blue, the more force necessary on the barbs. Blowing towards the ventral side (without the colors) shows that
one need stronger wind for separation the barbs. For more qualitative gross results, I have used an home desk 8 inch fan with
50 Hz 26 Watt. When the "eye" plane was parallel to the plane of the 3 rotating blades, about 2 m apart, the barbules of zone
5 were shaking but no separation occurred. When I approached the "eye", to about 1.3 m distance, the GoldenYellow zone 4 was
opened gradually. From about 1.0 to 0.6 m the RedBrown zone 3 was opened gradually. In about 0.5 to 0.3 m the Turquoise zone
2 was opened gradually. In about 0.25 m the DarkBlue color zone 1 was opened at once all along the way. Thereafter, when I
moved the "eye" far away of the wind, the barbs of the colored zones 1 to 5 were closed automatically. Approaching the "eye"
again, while its plane was not parallel to the plane of the rotating blades have caused to immediate separations of the barbs
along the colored zones of an "eye. Doing those qualitative experiments with "eyes" of different sizes reveal that the smaller
"eyes" need much stronger winds, for separation of the same colored zone. All those phenomena suggest looking for a connection
between the function of the train of the blue peacock and the variety kinds of winds this peacock may encountered in its wild
habitat.
The differences in the behavior of the barbs in each colored zone encouraged me to ask whether there
might be differences in their constructions. As we know, the barbs are "hooked" together by a zipper like mechanism. This
mechanism might be calibrated to the force of air blow necessary for the barbs to come detached. And indeed, using a magnifying
glass we can spot the differences in the density of the barbules found on different areas with different colors. In the Turquoise
zone 2 we can detect higher density of barbules than in the RedBrown zone 3, and the DarkBlue zone 1 holds higher density
than in zone 2. The particular structure of the barbs in each colored zone may be viewed when magnified by 100 or 400, using
a simple microscope. Then we realize that every color reveals different hooklets, teeth and structural shapes of barbules
in the different zones of the "eye of a peacock's train. One may see sketches of such barbules of a blue peacock's "eye" in
e.g. Fig. 26 of [Durrer, 1965]. This phenomena may be seen in other birds as well, see e.g. Fig. 255 of [Lucas, 1972] where
shown different structures of colored barbules in the pennaceous part of a humeral feather from a Bronze Turkey.
Thus, there are clearly different structures at different color zones of the "eye" of a peacock's
train. But, the colors themselves are due to the existence of layers of melanin, as pointed out in the summary of [Durrer,
1965] (p. 408) regarding the "eye": "The electronmicroscopical records of barbules with iridescent colours revealed that the
melanine granules are arranged in a space lattice. Interference of incident rays at the planes of the space lattice produce
the colour effects.
Therefore, there is an important question. If the main purpose of Nature was to create, along the
evolution, such a nice "eye" in the peacock's train for communications purposes, it was probably much simpler for Nature to
develop a one shape of barbules but with different spaces of melanin, which could give the desired colors. The fact that there
are different shapes of barbules in the different colored zones suggests that the aim of those zones of the "eye" is to fulfill
specific mechanical functions. The lattices of melanin may be used for reaching the desired mechanical functions. The above
mentioned behavior of each colored zone of the "eye", while gently separation of barbs by fingers or while blowing winds towards
the "eye", strengthen the hypothesis that the "eye" serves for certain mechanical functions. In order to find out the mechanical
functions of the eye" we have first to ask about the mechanical functions of tails and trains in pheasants.
3. Functions of tails and train in Pheasants
The Pheasants are bird family of the Galliformes order. Their source is in the tropical rainy jungles
of SouthEast of Asia. In those areas the strong monsoons winds govern the weather all over the year. The pheasants are big
birds in dimensions and weight. They are ground walker and lover of forests. In the wild they use to make short flies to the
branches of the trees for roosting and foraging for food. In the pheasants, the sexual dimorphism is highly developed. The
males are usually larger and heavier than the females. In most species of pheasants, the males have a long train in addition
to a short tail. The plumage of the pheasants is usually brilliant in many colors. See e.g. nice paintings by H. Jones in
[Johnsgard, 1986] and see nice photographs in INTERNET sites.
Generally, birds' tails are subject to gravitational and aerodynamic forces. Among the mechanical
functions of those tails I may mention: a) gravitational balance of the bird while it perches or jumps on branches. b) Air
brakes while landing. c) Aerodynamic stabilizer while the bird fly, perching or jump between branches. Let us discuss those
functions:
a) The gravitational balance: Let us imagine a heavy bird whose food is found on branches of trees
in such a jungle. It should have strong and not short legs in order to be able to jump from a branch to a branch while foraging
for plants and animals living on those trees. Thus the center of mass of its body, without the train, should be high above
the branch on which it stands. Lowering the tail downward may cause the center of mass to become on an imagine point within
the branch. This gives balance to the bird. Thus, the first function in that list is fulfilling. In pheasants this function
of the tails is even emphasized. From those paintings of Jones, we see that while a pheasant stands on a branch, its train
is inclined downward at an angle which depend on the position of the body. One may realize that the center of mass of the
pheasant body, without the train, is above the branch and somewhat forward, in the direction of the head. Without the train
(or the tail in the female), the pheasant may loose its gravitational balance and roll forwards. By position the train in
an inclined direction downward, the center of mass of the whole pheasant moves to an imagine point on the branch between the
legs of the pheasant. When the pheasant forage for food the situation is even worth. Since the legs of the pheasants are not
short, the pheasants need a long neck in order to reach the ground, or the level of the branch, while foraging. Sending the
head and the neck forwards cause also to the move of the center of mass forward. In the extreme case the plumb line from the
point of the center of mass may become out of the base" of the pheasant. This base is generally the surface below the fingers
of the legs. Thus, the body of the pheasant may loose balance and fall forward, as much as the problem with the Piazza tower.
In order to avoid this, the pheasant should change the position of the train, so that it may become more horizontally.
b) Air brakes while landing: This function is trivial and there is no need to explain it. I may only
mention here that airplanes use this function while landing, by lowering the edge of the wings.
c) Aerodynamic stabilizer: Let us discuss the aerodynamic functions of the pheasant's trains. These
trains may be used as an aerodynamic brake for stabilization while the pheasant is standing on a branch, and for some reason
it looses its balance and start rolling foreword or backward. During this motion there is a great velocity of air, relative
to the long train, and an aerodynamic resistance is developed on the train. As we know, the aerodynamic resistant on any body
is proportional to the square of the relative air-body velocity. Let us regard the branch as the axis of rotation while the
pheasant starts to roll forward or backwards. In this case the far parts of the train passes the greatest bows, thus they
have the greatest tangent velocity relative to the other parts of the pheasant. Therefore, the far parts of the train generate
the greatest aerodynamic resistance per a unit of surface. If there are palm-like at the edge of the feather's train, the
whole train may serve as an efficient sail. The longer the train the less total surface of the palms of the train is needed.
This argument is good for a long train of any pheasant. But it dose not explain yet the special "eyes" in the Pavo pheasant's
train. For understanding this we may ask about the specific needs of the blue peacock, in relation to the other pheasants.
According to [Johnsgard, 1986], the long tail (train) of the blue peacock, which it length is 1.4
- 1.6 m, comes in addition to it's short tail (0.40 - 0.45 m). The peahen has no train, but only a short tail (0.325 - 0.375
m). The short tail of those peafowl is used while they fly [Henry, 1971]. I may add here, by the way, that it seems to me
that the short tail of the male may also function as a biological instrument for erecting the train, using the physical lever
principle".
If we wish to understand the mechanical functions of the "eye", in the blue peacock's train, we have
first to find out the mechanical functions of the train itself. We may find those by looking for the differences in movements,
as well as in habits, between the peacock and the peahen.
Habitat and behavior of the Indian Peafowl.
As mentioned in some handbooks of birds, the source of the Indian Peafowl lives in special areas
near by rivers in a fairly thick jungles of India and Ceylon, e.g. [Baker, 1935; Whistler, 1941, Henry, 1971; Johnsgard, 1986].
I wish to quote here an important paragraph, from [Henry, 1971] regarding the Indian Peafowl: About sunset they fly into some
tall tree, which gives an uninterrupted view of countryside, to roost; not infrequently a top branch of a deed tree is chosen,
where they are exposed to all the vicissitudes of the weather. These they do not seem to mind, and the only effect a night
of rain seems to have upon them is that the cocks delay their descent in the morning until their trains have dried sufficiently
to be manageable. From this quotation one may conclude that not only the peacock but also the peahen was roosting on the top
branches of the tallest tree, in the wild tropical jungle. I argue that this was not the real situation. In addition, the
interpretation in the last sentence, regarding the reason to the delay of the peacock, does not seems to me as the main reason.
It is more reasonable that the main reason of the delay was that with the light of dawn the peacock would have started gathering
its exotic diet found at the canopy of the tropical rain forest, such as orchids and other epiphytes, and the insects feeding
on them. After several hours it become probably too sunny and those menu of the peacock were less available. Then he descent
to the peahen and young to supply them with the high quality food it had collected. It is reasonable to expect that each kind
of a pheasant may look after food at specific height levels of branch. The Indian Peafowl was known to reach the highest branches
in the tropical jungles of India and Ceylon. As described in [Johnsgard, 1986]. When roosting in forests, they select one
of the highest trees well out in the open." Up there it probably have found special plants and animals which may need to be
exposed to more sun then those on the lower branches. One may reasonably expect that large epiphytes and small animal, which
may live on those epiphytes, could be an example to the very fond food of the Indian Peafowl. I didn't find a review regarding
the diet of the wild peacock in those canopies. It was probably a hard task for the researchers, many years ago, to search
this point. Nowadays, it is recommended to do such a research, using radio transmitters and telephoto video camera, in the
remnants of the wild jungles of India and Seri-Lanka where there are yet some wild peafowls.
There are some facts, which may support my assumption that only the peacock was climbing to the top
of the tallest trees to roost there at night.
a) As noted in [Johnsgard, 1986] the incubation is by the female alone, almost 24 hours a day, for
28-30 days till the hatching. Thereafter, it took about two years till the young grow. During the first year the female protects
them while they are inside undergrowth, or on the low branches at nights and when there are monsoon floods. In addition, it
is accepted that the Pavo species is polygamous. Generally, the adult peacock (age 3 years or more) has a harem with tree
to five females. All this means a lot of work for the adult male in order to supply the peahens and young on ground with the
best food available.
b) The peacock have the known very loud and special voice, such as may-awe call [Johnsgard, 1986].
This call is not regarded as a territorial call since it is uttered before roosting, at the evening, and after roosting, early
in the morning. I may suggest that this call is a communicating call while departing between the male and the females, like
"good-bye", and before meeting again in the morning, like "hello". This call may be another indication that the male goes
far away of the females. The loud and special call might be for overcoming the large distances and the noises of the monsoon
winds within the jungle.
c) The peacock has a special hard finger in the back of its leg. The peahen generally does not have
it. The popular explanation to that finger is that its purpose is for fights between the males. I think this is a secondary
function. It seems to me more reasonable that the male uses that finger for digging inside the branches on the top of the
tallest trees in order to reach the special roots, seeds and the insects leaving inside the splits. The peahen may not need
such a finger since it lives most of the time on ground or on low branches.
All those points may indicate that the peacock was roosting and foraging for food on the top of the
tall trees, while the peahens were waiting on the ground or on low branches, for the supply of the special menu.
It is clear that the strongest winds in those jungles should be at the top of the tall trees. In
India and Seri-Lanka there are very strong winds due to the monsoons all over the year. While being there, it was very important
that the balance of the peacock's body should be automatically, no matter what kind of wind exist. Thus, if we wish to understand
the physical functions of the peacock's train we should look after the special kinds of winds the peacock has to stand.
I would expect the peacock to use the special train not only for balance while roosting and foraging
on the canopy of the jungles at night and early in the morning, but along the whole day. While the peacock is on the ground
he does not need the functions of the train for balancing. But he should carry after it. By erecting up the train it is easier
for him to carry it for long periods, since than there are no gravitational moments due to the long train. Only the weight
of the train is count. But this is very light. In addition, the peacock should open the train, and shake it from a side to
a side, probably in order to let all the barbs, along each feather of the train, to reset into the correct position.
I may expect also that a peacock is clever enough to see that the erection and shaking of the train
attracts attention. Thus, he may use it for impressing the peahen as well as for other communicating needs. However, these
options should be regarded as a secondary functions of the train, while the mechanical functions, mentioned above, seems to
be the primary reasons for the existence of the long train.
5. Qualitative physical computations
Let us discuss qualitatively the physics concerns the train of a peacock. In a zoo, and in the paintings
of Jones [Johnsgard , 1986], we see that while a peacock stands on a branch, its train declines downwards. It is reasonable
that while the peacock stands on a high branch at the canopy of the jungle, its train declines downwards as well. It is also
reasonable that for each peacock there is a certain angle beyond which the train can not be declined.
Suppose a non-turbulent planar wind, with an average velocity, Vb, blows towards the back of the
peacock while it stands on a branch. (We use here the suffix b for the body of the peacock). The wind pushes the body of the
peacock at a force Fb, which imposes an angular momentum Mb=FbLb about the branch, while the center of gravity of the peacock
(without the train) is at a distance Lb above the branch. Let Sb be the value of the surface of the back body of the peacock,
not including the train. The mass of air with density Q impinges on the body of the peacock at a specific interval of time
is QSbVb. For simplicity, let us assume that the air recoils elastically from the body. In this case the angular momentum
imparted to the body of the peacock is.
(1) Mb=2QSbLbVb2
Now let us consider the same simplified treatment to the train of the peacock, which is pointing
downwards. That planar wind exerts on it an angular momentum in the opposite direction to this exerted on the body. For simplicity,
let us regard the train as composing a long thin stick and a big palm at its edge, like a sail. The surface of the palm is
marked St and the distance of the palm from the branch is Lt. (I use here, the suffix t for train). The same simplified argumentation
leads to the equation of the angular momentum Mt of the train, about the branch exerted by the planar wind:
(2) Mt=2QStLtVt2
For balance, the two angular momentum must be equal: Mt=Mb. As long as the wind is planar we may
assume the velocities of the wind are the same for the body and the train, i.e. Vt=Vb. Therefore, from (1) and (2) we get
(3) St/Sb = Lb/Lt
This simple argumentation shows that, in principle, the condition for balance against planar winds
is that the ratios of the surface of the train to the surface of the body should be reciprocal to the ratios of their distances
from the branch. This conclusion is general, for any bird, not just for a peacock. The velocity of the wind is not included
in (3). Therefore, it holds no matter what is the strength of the wind, as long as it is planar. In open areas the wind might
be considered planar while regarding the scales of birds. For example, when a falcon stands on an electric wire, the position
of its tail maneuver according to weight balance and wind balance conditions. The balanced equation against planar winds should
be, in principle, according to (3).
One may see a very nice picture of a peacock with an open train in [Gilliard, 1967]. In that picture
the eyes" are ordered in several half circles, centered about the back of the peacock. There are no dimensions of the peacock
in that picture. However, according to the data given above I take, for simplicity, that picture as representative of a peacock
with 1.5 m long train. This assumption does not influence the theory. Using this scale I evaluate approximately the following
values: Lb= 0.24 m, Lt = 1.2 m and Sb= 145 cm2. Using (3) shows that these values lead to a value of the surface
of the train needed for planar winds: St=29 cm2 . But, this is about the surface of one big "eye at the outer circle
of "eyes". Therefore, in principle, if the only problem of the peacock was to balance against planar winds, it might be sufficient
to have several long feathers only, with surface of a palm as big as one such "eye. But, there are about 150 feathers (each
eye" is on the edge of one feather). All those feathers might be needed mainly for lowering the center of mass in order to
bring it onto the branch, which the peacock stand on, while the train is pointing downwards. This is due to the weight balance.
But, so many long feathers in the train, with total surface larger, say, 100 times than needed for wind balance, may cause
lot of troubles to the peacock even weak planar wind. There is a crucial need for the peacock to reduce the total surface
of the train when there is a wind beyond a specific strength. Closing the train may be too slowly and may also be insufficient
for winds above a certain strength.. There is a better way as I show in the following general case.
In the general case of turbulent wind, one should consider the balance equation due to the contribution
from each feather. The angular momentum of the train in the general case, Mtg, when there are various lengths and various
wind velocities, is:
Mtg = 2QiSiLiVi2
while the summation is over all the feathers i with surface Si and length Li which are impinged by
local wind velocity Vi.
In a turbulent wind the values of the air densities Q are local as well. But the differences are
small and we take in (4), for simplicity, one value Q for all the feathers. As in the former case, for balance we demand Mtg=Mb.
From (1) and (4) we get:
(5) SbLb =iSiLi(Vi 2 / Vb 2 )
On the left hand of (5) all the parameters are constant. On the right hand all the parameters might
be non-constant due to change of positions of the feathers as well as the local relative velocities of the turbulent winds,
which may differ in time and space. If the wind is planar than Vi=Vb and (5) becomes (6) SbLb = iSiLi.
Let us try to evaluate the value of the right hand of (6), for a typical train of an adult peacock.
Let us consider, for a gross computation, that there are 150 "eyes in an open train consisting of 15 bows, with 10 equal "eyes"
in each bow. In addition suppose the radii of the bows goes from 10 cm to 150 cm, in intervals of 10 cm, and the surfaces
of the "eyes" in those bows goes from 1 to 15 cm2. Under those data, the right hand of (6) gives, for the train,
124,000 cm3. At the same level of gross computation we may consider the surface of the body of the peacock, Sb,
to be 1000 cm2 and the distance to the center of mass, Lb, to be 40 cm. Thus, on the left hand of (6) the value
accepted for the body is 40,000 cm3 , which is about third of the value computed for the train in the right hand
of (6). Therefore, for balance during changeable winds the train should be opened and closed according to the total moment
it exerts on the peacock. Those changes in the positions of the train may be done easily if the changes in the local velocities
are graduate and predictable, as might be the case in open areas or on the low branches of trees in a jungle, where one finds
pheasants with various kinds of long trains [Johnsgard 1986]. But, the turbulent monsoon winds at the highest branches of
the tropical rain jungle are strong, changed abruptly and cannot be predicted. In those places there is no time for the peacock
to close and open the train. There should be some automatic mechanism to control the total sum out of the surfaces of the
feathers, in the long train, to be adapted instantaneously to the changing conditions so that (5) may hold and the peacock's
balance conserved. As I show in the following, the nice "eye" in each of such long feather is the most wonderful physical
mechanism for controlling those critical situations.
6. Physical functions of the "eye
As we see above, there is a crucial need for the peacock to reduce the total surface of the train
when the strength of the wind is beyond some level. Closing the train may not be enough above a certain strength and there
is needed an efficient mechanism. One of the ways for controlling the balance against strong monsoon winds on a peacock's
train is by letting the barbs of all the "eyes" of the outer circles, to be separated. So they are not count in the total
surface St of the train since the wind passes through them
As we see in that picture of the peacock's train, there are bows of half circles of "eyes". The radius
of the inner circle of eyes" is about 10 cm. The size of these "eyes" is much smaller than the size of the "eyes" at the outer
circle. Some of the small "eyes" in the inner circle have about 3 cm2. According to (3), for balancing with a rod
of the length of this radius, 10 cm, there is a need for a palm of 200 cm2. That means about 67 of such small "eyes".
We see in the picture that there are less than 15 "eyes" in that circle. So, for balancing the wind in the planar case there
should be "eyes" at circles of medium radius that their barbs are not separated. This argument is good when there are only
two options for each "eye: totally open or totally closed. But there is yet another, much more useful, possibility for overcoming
the unbalanced problem. This is when in many "eyes" only part of the "eye", say the DarkBlue zone 1, remain closed. This zone
has lower surface than the total "eye". For comparison, in one typical "eye" I have measured qualitatively the surfaces of
color zones. The gross values I got are:
Zone 1: 204 ± 20 mm2
Zones 1+2: 409 ± 30 mm2
Zones 1+2+3: 1085 ± 50 mm2
Although these are gross measurements, using a millimeter page, it shows qualitatively that the surface
of the DarkBlue color zone 1 is about fifth of the surface of the whole "eye", while the barbules are totally closed. Thus,
according to (2), if only the surfaces of zones 1 are closed the train may reach balance even against a wind which its velocity
is 2.23 (square root of 5) times stronger than when the whole "eyes" are closed.
All the calculations, so far, were regarded to a planar linear wind. The meaning of linear here is
that the wind arrives from one direction only. This linearity may exist in open areas or among trees when the wind is quiet
weak. But at the upper branches of the thick forest, as in a tropical jungle, monsoon winds may be relatively strong and very
turbulent. When a linear wind goes through any physical body there are generated vortices of turbulent air behind the body.
The width of these vortices depends on the width of the body, the strength of the wind, the density of air, etc. One may see
such vortices in a stream of water while there is a body inside the stream. See e.g. p. 142 in [Hertel, 1966]. One may also
see this phenomena at a bathtub. Up there, at the top of jungle trees, any strong wind may create a lot of vortices in all
directions and sizes. Some of them may be as small as 1 cm width or less, other may be as large as 10 cm width or more. This
depends on the width of the branches and many other factors, according to the theories on turbulent fluid dynamics. When a
peacock stands on a branch at the top of a tall tree, while there is a strong wind, it is possible that one part of the peacock
may feel a totally different kind of wind relative to another part. It is also possible that a one feather of the train may
feel much stronger wind, and from different direction, relative to adjacent feather in that train. A usual pheasant with a
"simple" long train without "eyes", can do almost nothing about such a situation. It will probably get out of balance and
fall down of the branch. Here enters the main mechanical function of an "eye" of a peacock's train. As we saw above in the
experiments using a ventilator, the "eye" is very sensitive to winds that are not normal to its plane, i.e. "side winds".
In that case the barbs are separated immediately even in weak winds. These "side winds" make the same mechanical action as
I did while separating two barbs of an "eye" using two fingers. Vortices mean a lot of side winds on the feathers. To prevent
unbalanced forces, on the different feathers of the train, it is important to allow the palm-shaped "eye" to be opened instantaneously
and with the proper amount of opening. Yet, it is very important to keep some of the surface of the "eyes" closed, i.e. no
separation of all colored barbules. This is because the train should yet be able to balance frontal or backward winds in the
manner described above. It is also important to note the special V-shaped at the top of the longest feathers in the peacock's
train as in p. 80 of [Simpson, 1986] or p. 271 of [Johnsgard, 1986]. As a side-wind hits this V-shaped, it may twist the feather
towards the direction of the wind. This may cause the skin near the base of this feather to feel the twisting. Thus, the whole
train may rotate towards the direction of the side wind so that the wind will be frontal to the train and the balancing procedure,
mentioned above, may be applied.
As a result of those qualitative arguments I arrive at an idea that the train of the blue peacock
may use as a very sensitive balanced instrument against winds of variety strengths, directions and periods of times. This
instrument should be able to balance the blue peacock while it forages or roosts on a high branch when there could be all
sorts of winds: linear, circular or turbulent winds coming in front, behind or in angle to the blue peacock at short or long
blows, etc. What also important is the automatic resetting of the instrument to the normal situation, when all the barbs of
the "eye" are closed. This is necessary for controlling the next blow of wind that might be totally different than the former
one. We saw in the above simple experiments that this resetting is achieved automatically in the "eye" of the blue peacock's
train.
Thus, among the mechanical functions of a bird's tail, mentioned above, I may emphasize here the
stabilization function of the train while the pheasant is perching. This stabilization is not the same as stabilization during
flight. Therefore, while the peacock fly, "the tail is widely spread, fan-wise, with the train (comprising the rump-feathers
and upper tail-coverts) compressed into a narrow bundle" [Henry, 1971].
7. Structure, materials and colors in the "eye
One of the most interesting questions is what is the connection between the mechanical functions
of the peacock's train, mentioned above, and the special colors of the "eyes" in such a rain. An optional answer is as follows.
As we see, the barbules of the "eye" should be opened at specific distances from its center, due to specific strengths of
winds in specific directions. Therefore, there should be the correct structures of the barbules in specific distances from
the center of the "eye, so as to enable the separations of barbules in the defined mechanical conditions. The barbules should
have the correct mechanical properties, like density, strength, flexibility, etc. In order to achieve those properties, the
covering of the barbules should be in specific stiffness according to their mechanical functions. We shall see in the following
that this is indeed the situation.
In Tafel VII (last page) of [Durrer, 1965] we see clearly the different structures of layers of granules
of melanin at color zones in the "eye" of a peacock's train. The 40,000 magnification, using an electron microscope, reveals
the specific combinations of spacing between layers, in a barbule, needed for "creating" a specific physical color. In Tables
2 and 3 of that article there are the dimensions of the lattice of granules in each colored zone. From those three tables
I summarize in Table 1 here data relating to the zone numbers I have noted above.
Table 1: Dimensions of granule lattices of colored zones in the "blue peacock's "eye .
( = wavelength ; N = no.
of layers; d = measured spacing between layers).
Zone Color (mµ) N 3d (mµ)
±
------------------------------- ---------------------------------------------
1 DarkBlue 450 - 470 9 - 11 480 ± 18
2 Turquoise 490 - 510 9 - 10 510 ± 15
3 RedBrown 590 - 620 5 - 7 630 ± 21
4 GoldenYellow 570 - 600 4 - 6 624 ± 40
5 Green-Red 500 - 630 3 - 6 630 ± 45
----------------------------------------------------------------------------
Note: the value 3d influences the appearance of the color due to interference from the
lattice of granules imbedded within a keratin with refractive index 1.5 (See
Fig. 38 there).
From Table 1 we see that there is a correlation between the colored zones and the quantity and density
of melanin layers imbedded within the barbules of those zones. These contribute to the mechanical properties of the barbules.
One may learn it from e.g. p. 393 of [Lucas 1972]: "Melanin also has the effect of making a feather denser and more resistance
to wear and photochemical changes. This property of melanin has long been known from gross observations and has recently been
supported in studies with an electron microscope. From his observations, Carr (1957: 161) gained the impression ' that the
particles arranged in lengthwise rows, contributed quite notably to the bulk and strength of the feather .
So, the bulk and strength of the barbules at the colored zones are influenced by the layers of melanin
imbedded within the keratin substance of those barbules.
Let us see now how all this influence the characteristics of the separations between the barbs in
the "eye" of the blue peacock train. In Fig. 73 of [Hertel , 1966] we see a cross section through three barbs of a pheasant
feather, with the shape C, while they are anchored together at the top of the C by barbules. As explained in p. 57 of that
book of Hertel, the connections between two barbules of neighbor C-shaped barbs depends on the bending strength and resistance
of those C-shaped barbs. It is reasonable that when the strength is higher the connection is stronger. Therefore, we may expect
that when the melanin layers within specific barbs are denser, they have more strength and therefore the connections between
the barbules of those barbs are stronger. This is with regard to the barbs. Let us see what happens with the barbules. As
we may understand from the above mentioned sketches of the barbules in Fig. 26 of [Durrer , 1965], one may look, in principle,
upon a barbule with a hooklet at its upper edge, as upon a metal spring with a hook. For simplifying the explanation, let
us regard this hook as if it is caught within a small ring. If the spring is very flexible, it will be easy to eject the hook
from the ring. You only have to bend or push it a little bit and the spring vibrates till the hook is out of the ring. But,
if the spring is less flexible, it will be harder to eject the hook. In principle, the same might have happen in the barbules.
Where the layers of melanin are denser, in a specific part of the barbule, this part is more strengthen, i.e. it is less flexible.
Thus it is harder to eject the hooklets of a barbule with higher density of melanin layers. This may explain why it was more
difficult to separate, e.g., the barbules in zone 2 (turquoise) than those in zone 3 (RedBrown) of a peacock's "eye". From
the last column of Table 1 we see that in zone 2 the spacing between two layers of melanin is less then in zone 3. As I have
pointed out above, the barbules in zone 1 ( DarkBlue) was resistant to opening but when it was starting to be opened, it was
abruptly opened all along the way. The bent shape of the smaller barbules in this zone and the tiny hooklet at the upper edge,
as seen in Fig. 26 of [Durrer, 1965], may explain this phenomena, while considering the example of the metal spring given
above. As we see in Table 1, in this zone 1 the spacing between the melanin layers is the smallest. Therefore, the color is
of shorter wavelength then in zones 2, 3, 4 and 5.
Thus, we have a reasonable physical explanation to the correlation between the colors of the barbs
in the "eye" and their separation characteristics under mechanical conditions. Now, that we understand all this, we may ask
about the purpose of the pattern of the whole eye. The semi-symmetrical patterns of each zone in the "eye", as described above,
may show about the needs to open the zone at specific parts, due to specific kind of winds, so that the whole feather will
not be twisted or bent beyond certain levels. The symmetrical pattern of the whole train of the peacock may serve to stabilize
aerodynamic balance, against all kinds of wind form any direction at any period of time, without loosing the weight balance
achieved by the train.
Thus, we have an optional physical explanation to the extraordinary pattern in the "eye" of the blue
peacock's train. This "eye" may serve as a special mean to let the train stabilizing the blue peacock while there are variety
kinds of winds at different strengths, directions and frequencies of blows. It might be expected that such a train was developed
along the evolution while the peacock was climbing higher and higher up to the canopy of the tallest trees, in order to find
more and more delicate food, in spite of the monsoon winds in the tropical rainy jungle.
8. Conclusion
In this article I suggest a physical explanation to the "eye" at a feather of a blue peacock's train.
I show that the "eye" may use as a device for controlling the strength of wind passing through the palm-like at the edge of
the feather's train. Each colored zone in the "eye" may serve as controller for allowing a specific strength of wind to pass
through it. The closer the shade to DarkBlue, the more strength of wind is necessary to separate the barbs in the "eye". The
control mechanism, suggested here, is by letting a separation of the barbules in that "eye, by the right amount, under all
kinds of winds. This includes turbulent winds coming from various directions and strengths while they may be locally changed
abruptly on each feather. All this might happen when the blue peacock is foraging for special food on the highest branches
of the tallest trees in the tropical rainy jungle in India and Ceylon, where strong monsoon winds govern the weather all over
the year. I argue that the blue peacock was actually collecting delicate food, on the canopy of those jungles, for the peahens
and their young, who were waiting on the ground or on low branches in cases of floods.
I have not dealt here with the other species of the Pavo genus, i.e. the Green Peafowl and the Afropavo.
According to [Johnsgard ,1986] the Green Peafowl habitats are Bangladesh to Indochina the Island of Java, in lowland and moderate
altitude forests, and in open areas from the plains to about 900 m. As one may see in the maps of winds, the winds in those
areas are not strong, in comparison to the monsoon winds in India and Seri-Lanka. Therefore, I am not surprise that the colors
of the Green Peafowl are closer to the green than to the blue. This is in accordance with my above conclusion that the BlueDark
barbs need more strength of wind than the turquoise and longer wavelength colors, in order to separate between the barbs.
The other species is even more convincing. The main colors of the Afropavo are even of longer wavelength than those of the
Green Peafowl. This hints that the Afropavo habitats are in areas where the winds are much weakest than in the areas of the
Green Peafowl. And indeed, the Afropavo was discovered not long ago in Kongo, and both the male and the female do not have
a long train. This indicates that this species do not forage for food at places of strong winds. From the wind maps we see
that the winds in Kongo are much weakest than the winds in the southeast of Asia, the source wild land of the pheasants with
long trains.
I recommend checking my physical explanation more deeply. In a deep research one may have to follow
blue peacocks, peahens and other pheasants in their natural habitat. By using radio transmitters and telephoto video camera,
one may research the specific food and behavior of each species and sex in the canopy of the jungles and on the ground. In
addition, one may measure the turbulent winds at different heights in those jungles and compare all these data to the mechanical
functions of the trains, and in particular with the extraordinary functions of the blue peacock's "eye.
Acknowledgments
I wish to thank Dr. Ilana Steinhorn for her contribution to this article. I also wish to thank Dr.
Neta Maoz, from the Wiezmann Institute of Science in Rehovot, Israel, for encouraging me along the last 10 years in developing
the new theory.
References
1. Seife, C., New Scientist, 6 Mar. 1999, p. 10.
2. Sword, G.A., Nature , Vol. 397, 21 Jan. 1999, p. 217.
3. Vogel, S., Physics Today, Nov. 1998, pp. 22-27.
4. Johnsgard, P. A. (1986). The pheasants of the world. Oxford University Press.
5. Welty, J.C. (1982). The Life of Birds, (third edition) Saunder College Publishing.
6. Lucas, A.M. & Stettenheim, P.R. (1972). Avian Anatomy Integument, Part II,
U.S. Government Printing Office, Washington, D.C.
7. Henry G.M. (1971), A Guide to the Birds of Ceylon, Oxford University Press.
8. Gilliard, E.T. (1967), Living Birds of the World Doubleday & Company, Inc., Garoen City, New
York.
9. Hertel, H. (1966). Structure Form and Movement. Reinhold Publishing corporation, New York.
10. Durrer, H. (1965), Bau und Bildung der Augfeder des Pfaus (Pavo cristatus L.), Rev. Suisse de
Zool. 72. pp. 263-411.
11. Whistler, H. (1941). Popular handbook of Indian birds, Gurmy and .Jackson, London.
12. Baker, E. C. S. (1935). The nidification of birds of the Indian Empire Vol. IV. Taylor and Francis,
London.
13. Petrie M. and T. Halliday, Behavioral Ecology and Sociobiology 35(3), pp. 213-217 9/1994.
| Fig.1 Schematic picture of the "eye" with 5 zones |
|
|
